Orgánulos celulares: mitocondrias, cloroplastos, retículo endoplasmático, complejo de Golgi, lisosomas y vacuolas.
Las células eucariotas presentan un complejo sistema de membranas interno que llega a ocupar la mitad de la célula, el llamado sistema endomembranoso, que divide el contenido celular en compartimentos. En cada uno de ellos se realiza un tipo de reacciones bioquímicas, evitándose así que unas interfieran con otras. Estos compartimentos son: el retículo endoplasmático, el aparato de Golgi, las vacuolas, los lisosomas y los peroxisomas. Además, hay dos tipos de compartimentos, no relacionados estructuralmente con el sistema endomembranoso, que poseen una doble membrana y que son orgánulos productores de energía. Son los cloroplastos y las mitocondrias.
MITOCONDRIAS
Las mitocondrias son los orgánulos de las células eucariotas que se encargan de la obtención de energía mediante la respiración celular, un proceso de oxidación en el que intervienen unas enzimas denominadas ATP-sintetasas. La energía obtenida se guarda en forma de ATP. Aparecen en grandes cantidades en el citoplasma de todas las células eucariotas, tanto de animales como de vegetales, siendo especialmente abundantes en aquellas que por su actividad poseen una elevada demanda de energía bioquímica (ATP). El conjunto de mitocondrias de una célula se denomina condrioma.
Estructura de las mitocondrias
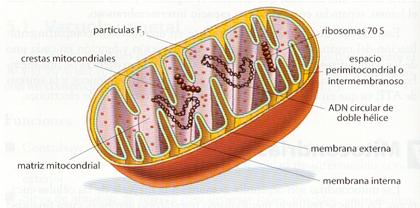
Las mitocondrias son orgánulos polimorfos, pudiendo variar desde formas esféricas hasta alargadas a modo de bastoncillo. Sus dimensiones oscilan entre 1 µ y 4 µ de longitud y 0,3 µ y 0,8 µ de anchura. Presentan una doble membrana: una membrana mitocondrial externa lisa y una membrana mitocondrial interna con numerosos repliegues internos, denominados crestas mitocondriales. Estas membranas originan dos compartimentos: el espacio intermembranoso, entre las dos membranas, y la matriz, espacio delimitado por la membrana interna.
La membrana mitocondrial externa posee un gran número de proteínas transmembranosas que actúan como canales de penetración. A continuación, se encuentra el espacio intermembranoso, de contenido similar al del citosol. La membrana mitocondrial interna presenta repliegues o crestas que incrementan su superficie y, por tanto, su capacidad metabolizadora. Es bastante impermeable, y presenta un gran número de proteínas de membrana que desarrollan una amplia gama de funciones, destacando las permeasas, los componentes de las cadenas moleculares transportadoras de electrones, entre los que destacan los citocromos, y los complejos enzimáticos formadores de ATP, denominados ATP-sintetasas. Entre sus lípidos de membrana no aparece el colesterol, al igual que en la membrana plasmática bacteriana.
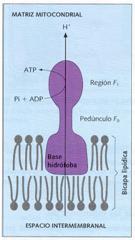
Las ATP-sintetasas están constituidas por tres partes: una base hidrófoba, que se ancla en la membrana, un pedúnculo o región F o, y una esfera de unos 90 Å de diámetro, o región F 1, que es donde se catalizan las reacciones de síntesis de ATP.
En la matriz mitocondrial existe un medio interno rico en enzimas y en el que se lleva a cabo un gran número de reacciones bioquímicas. Esta cámara interna presenta ribosomas mitocondriales o mitorribosomas, similares a los bacterianos, y varias moléculas de ADN mitocondrial, circular y de doble hebra como los bacterianos.
|
|
|
Estructura de una mitocondría
|
ATP-sintetasa
|
Función de las mitocondrias
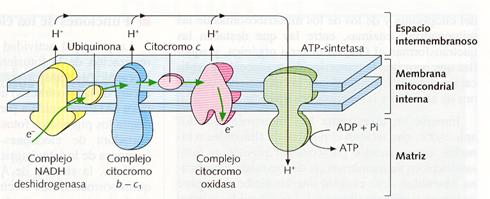
Su actividad principal es la respiración mitocondrial, cuya última etapa es la cadena respiratoria que se realiza en la membrana interna. En esta se dan los NADH y los FADH2 procedentes de otras vías metabólicas, obteniéndose energía que se almacena en moléculas de ATP. La síntesis de ATP por la ATP-sintetasa se explica mediante quimioósmosis: El NADH y el FADH2 liberan protones (H+) y electrones (e-). Éstos, al pasar por la cadena de proteínas transportadora de electrones que hay en las crestas mitocondriales, ceden energía, que es utilizada para bombear protones H+ fuera de la matriz. Éstos regresan a la matriz a través de las ATP-sintetasas donde la energía del gradiente es utilizada para formar ATP.
Sector de la membrana interna mitocondrial mostrando la secuencia de proteínas de la cadena respiratoria y la ATP-sintetasa que realizan la fosforilación oxidativa que conduce a la formación de moléculas de ATP
En la matriz mitocondrial se realizan otras vías metabólicas importantes; las principales son:
- El ciclo de Krebs.
- La b-oxidación de los ácidos grasos.
- La biosíntesis de proteínas en los ribosomas.
- La duplicación del ADN mitocondrial.
CLOROPLASTOS
Los cloroplastos son unos orgánulos típicos de las células vegetales que poseen clorofila, por lo que pueden realizar la fotosíntesis, proceso en el que se transforma la energía luminosa en energía química contenida en la molécula de ATP. Por ello, al igual que las mitocondrias, los cloroplastos son orgánulos productores de energía.
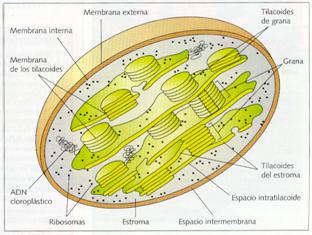
Estructura de los cloroplastos
Los cloroplastos son polimorfos y de color verde debido a la presencia del pigmento clorofila. En las algas, las formas son muy diversas; por ejemplo, en el alga Spirogyra sólo hay dos y tienen forma de cinta en espiral. En las plantas superiores, la forma más abundante es la de disco lenticular, aunque también los hay ovoides y esféricos. Miden entre 3 y 19 µ. de diámetro mayor y de 1 y 2 µ de diámetro menor, y suele haber entre 20 y 40 por célula.
Presentan una envoltura constituida por una doble membrana: una membrana plastidial externa y una membrana plastidial interna. La externa es muy permeable, mientras que la interna es casi impermeable, por lo que posee una gran cantidad de permeasas, denominadas proteínas translocadoras. Ambas membranas carecen de clorofila y entre sus lípidos, al igual que en las mitocondrias, no está el colesterol.
En el interior, delimitada por la membrana plastidial interna, hay una cámara que contiene un medio interno, denominado estroma, que posee un elevado número de componentes. Éstos son: ADN plastidial, circular y de doble hélice, como el de las bacterias; plastorribosomas, distintos de los ribosomas del citoplasma y de los mitorribosomas de las mitocondrias; enzimas, entre las que destacan las que transforman el CO2 en materia orgánica, y aquellas que permiten la transcripción, traducción y replicación del ADN; y finalmente, las inclusiones de granos de almidón y las inclusiones lipídicas.
Inmerso en el estroma hay numerosos sáculos aplastados, que reciben el nombre de tilacoides o lamelas, caracterizados por contener pigmentos fotosintéticos en su membrana, la denominada membrana tilacoidal, cuya cavidad interior recibe el nombre de lumen o espacio tilacoidal. Los tilacoides pueden extenderse por todo el estroma, por lo que reciben el nombre de tilacoides de estroma, o pueden ser pequeños, tener forma de disco y presentarse apilados como montones de monedas, los denominados tilacoides de grana, ya que cada montón recibe el nombre de grana. En las membranas de los grana se ubican los sistemas enzimáticos encargados de captar la energía luminosa, efectuar el transporte de electrones y formar ATP.
Estructura del cloroplasto.
Funciones de los cloroplastos
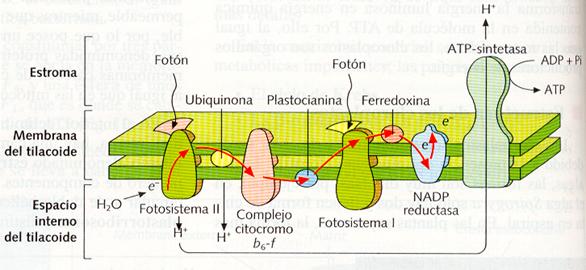
La principal actividad de los cloroplastos es la realización de la fotosíntesis, en la que la materia inorgánica es transformada en materia orgánica (fase oscura de la fotosíntesis) utilizando la energía bioquímica (ATP) obtenida a partir de la energía solar, mediante los pigmentos fotosintéticos y la cadena transportadora de electrones de los tilacoides (fase luminosa de la fotosíntesis). Al igual que en las mitocondrias, la síntesis de ATP se realiza mediante la quimioósmosis. Los electrones excitados por la luz, es decir, ricos en energía, al recorrer la cadena de proteínas situadas en la membrana de los tilacoides (cadena transportadora de electrones), van cediendo su energía, que es utilizada para bombear H+ hacia el interior de los tilacoides. Esto origina un gradiente químico de H+, cuya energía es utilizada por las ATP-sintetasas para la formación de ATP.
Sector de la membrana de un tilacoide, mostrando las proteínas que realizan el transporte de electrones y a la ATP-sintetasa.
Otras vías metabólicas, que se realizan en el estroma, son la biosíntesis de proteínas y la replicación del ADN.
RETICULO ENDOPLASMATICO
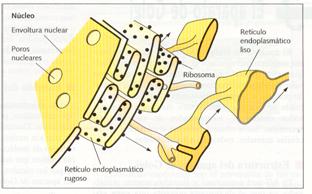
El retículo endoplasmático es un sistema membranoso compuesto por una red de sáculos aplastados o cisternas, sáculos globosos o vesículas, y túbulos sinuosos, que se extienden por todo el citoplasma y que se halla en comunicación con la membrana nuclear externa. Este sistema constituye un único compartimento con un espacio interno que recibe el nombre de lumen. Se distinguen dos clases de retículo endoplasmático: el retículo endoplasmático rugoso o granular (RER), antes denominado ergastoplasma, que posee ribosomas en su cara externa, la llamada cara citoplasmática y el retículo endoplasmático liso o agranular (REL), que carece de ribosomas.
Estructura del retículo endoplasmático rugoso
El retículo endoplasmático rugoso está formado por sáculos aplastados comunicados entre sí. Además, puede presentar vesículas. Se encuentra comunicado con el retículo endoplasmático liso y con la membrana externa de la envoltura nuclear.
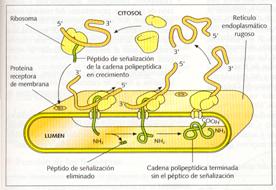
De hecho, puede considerarse que la envoltura nuclear es la parte del RER que separa el núcleo del citoplasma. Sus membranas son algo más delgadas que las plasmáticas (50 a 60 Å), y presentan proteínas encargadas de fijar los ribosomas, las riboforinas, y otras que actúan como canales de penetración de las proteínas sintetizadas por estos ribosomas.
Función del retículo endoplasmático rugoso
La función básica del retículo endoplasmático rugoso es la síntesis de proteínas mediante los ribosomas de su membrana, su introducción en el lumen, la glucosilación de las proteínas (que se completará en el aparato de Golgi), y su transporte hacia los orgánulos, donde son utilizadas para constituir membranas.
Este transporte se realiza en el interior de vesículas que se producen en la membrana del retículo endoplasmático rugoso.
Retículo endoplasmático rugoso y su relación con la envoltura nuclear y el retículo endoplasmático liso.
| |
Síntesis de proteínas en la membrana del retículo endoplasmático rugoso.
| |
Estructura del retículo endoplasmático liso
El retículo endoplasmático liso está constituido por una red de tubulos unidos al retículo endoplasmático rugoso y que se expande por todo el citoplasma. La membrana del retículo endoplasmático liso posee gran cantidad de enzimas cuya principal actividad es la síntesis de lípidos.
Función del retículo endoplasmático liso
En el REL se sintetizan casi todos los lípidos constituyentes de las membranas: colesterol, fosfolípidos, glucolípidos, etc. Sólo los ácidos grasos se sintetizan en el citosol. Estos lípidos se construyen en el lado citoplasmático de la membrana, desde donde se difunden hacia el interior del retículo liso. Estos lípidos se transportan a otros orgánulos, mediante proteínas de transferencia o por vesículas producidas por medio de las redes de clatrina por gemación. Es decir, intervienen en la síntesis, almacenamiento y transporte de lípidos.
Participa también en procesos de detoxificación, siendo capaz de metabolizar sustancias tóxicas y convertirlas en productos eliminables por las células.
Por último, interviene en la conducción de impulsos nerviosos para la contracción del músculo estriado.
COMPLEJO DE GOLGI
El aparato de Golgi (AG) forma parte del sistema membranoso celular. Fue descubierto por C. Golgi en 1898. Está formado por uno o varios dictiosomas (agrupación en paralelo de cuatro a ocho sáculos discoidales denominados cisternas), acompañados de vesículas de secreción. Suele situarse próximo al núcleo, y, en las células animales, rodeando a los centríolos.
Estructura del aparato de Golgi.
El AG está estructural y fisiológicamente polarizado, ya que el dictiosoma presenta una cara cis, próxima al RER, generalmente convexa, constituida por sáculos de menor diámetro y de membrana más fina, y una cara trans, próxima a la membrana citoplasmática, generalmente cóncava, y caracterizada por presentar cisternas de gran tamaño, de membrana más gruesa y de aspecto reticular. La cara cis o de formación recibe vesículas (vesículas de transición) procedentes de la envoltura nuclear y del retículo endoplasmático, que alimentan al aparato de Golgi. El contenido molecular del dictiosoma va avanzando hacia la cara trans o de maduración. Esta progresión se realiza de cisterna a cisterna, mediante pequeñas vesículas (vesículas intercisternas) y, una vez que llega a la cara trans, es concentrado y acumulado en el interior de unas vesículas mucho mayores que las anteriores (vesículas de secreción). Éstas pueden actuar como lisosomas si contienen enzimas digestivas o pueden dirigirse hacia la membrana plasmática donde pueden verter su contenido al medio externo (exocitosis) y además soldarse a ella y, así, hacerla crecer o regenerarse.
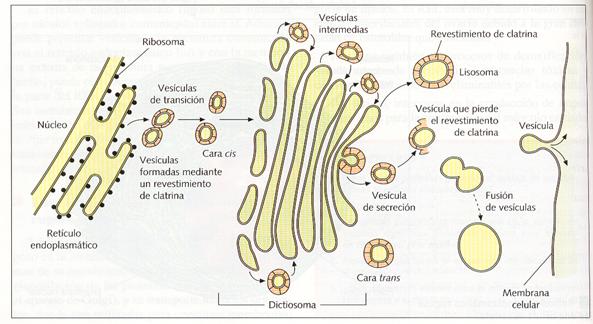
Origen del aparato de Golgi y los lisosomas.
Función del aparato de Golgi
Las membranas del AG son ricas en proteínas tipo de enzimático, por lo que pueden realizar múltiples funciones. Desempeña el papel de organizador de la circulación molecular de la célula. Por él pasan gran número de moléculas procedentes del retículo endoplasmático que sufren una maduración en su recorrido por los sáculos del dictiosoma.
Entre las funciones del aparato de Golgi destacan:
- Transporte, maduración, acumulación y secreción de proteínas procedentes del retículo endoplasmático. Así, muchas proteínas varían su estructura o alteran las secuencias de aminoácidos haciéndose activas. Posteriormente son concentradas y pasan al interior de vesículas de secreción.
- Glucosilación de lípidos y proteínas, mediante la unión a éstos de cadenas de oligosacáridos, dando lugar a glucolípidos o glucoproteínas de membrana, o de secreción.
- Síntesis de proteoglucanos (mucopolisacáridos), que son parte esencial de la matriz extracelular, y de los glúcidos constitutivos de la pared celular vegetal (pectina, hemicelulosa y celulosa).
LISOSOMAS
Los lisosomas son vesículas procedentes del aparato de Golgi que contienen enzimas digestivas. Éstas son hidrolasas ácidas (fosfatasa ácida, glucosidasas, lipasas, proteasa, ADNasa, etc.) que se forman en el retículo endoplasmático rugoso, pasan al aparato de Golgi, en donde se activan y se concentran, y que se acumulan en el interior de los lisosomas.
Estructura de los lisosomas
Los lisosomas poseen una membrana plasmática con las proteínas de su cara interna muy glucosiladas. Estas glucoproteínas impiden que las enzimas hidrolasas ataquen a la propia membrana del lisosoma.
Función de los lisosomas
Los lisosomas realizan la digestión de materia orgánica. La principal enzima digestiva es la fosfatasa ácida, capaz de romper los enlaces fosfoestéricos y liberar grupos fosfato. Para el buen funcionamiento de estas enzimas, los lisosomas necesitan mantener un pH entre 3 y 6; por tanto, introducen protones (H+) en su interior mediante gasto de ATP.
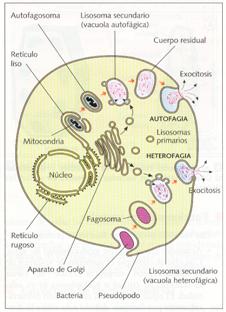
La digestión puede ser extracelular, cuando los lisosomas vierten sus enzimas al exterior, o intracelular, cuando se unen a una vacuola que contiene la materia a digerir. Se utiliza el término de lisosoma primario para referirse a los que sólo poseen en su interior enzimas digestivas y el término lisosoma secundario para aquellos que, por haberse unido a una vacuola con materia orgánica, contienen sustratos en vía de digestión. Los lisosomas secundarios reciben el nombre de vacuolas digestivas o heterofágicas, cuando el sustrato procede del exterior por fagocitosis o pinocitosis, o de vacuolas autofágicas, cuando procede del interior, por ejemplo, con moléculas u orgánulos propios, que previamente han sido envueltos por cisternas del retículo endoplasmático.
Esquema de la acción de los lisosomas en heterofagia y en autofagia.
Lisosomas especiales son el acrosoma de los espermatozoides y los granos de aleurona de las semillas. El acrosoma es un lisosoma primario en el que se almacenan enzimas capaces de digerir las membranas foliculares del óvulo, para permitir el paso del espermatozoide y la fecundación. Los granos de aleurona son lisosomas secundarios en donde se almacenan proteínas que, debido a la pérdida de agua, se encuentran en estado cristalino, hasta que al plantarse y absorberse agua se activan las enzimas y se inicia la digestión de las mismas, con lo que empieza la germinación de la semilla.
Los peroxisomas y los glioxisomas.
Los peroxisomas son orgánulos parecidos a los lisosomas, pero en vez de contener enzimas hidrolasas contienen enzimas oxidasas, entre las que destacan la peroxidasa y la catalasa.
Los glioxisomas son una clase de peroxisomas que sólo se encuentran en las células de los vegetales.
Una función de los peroxisomas es la detoxificación, proceso que elimina sustancias tóxicas oxidándolas. Así se degradan, en las células hepáticas, el etanol de las bebidas alcohólicas y otras sustancias tóxicas como el metanol, el ácido fórmico, etc.
VACUOLAS
Las vacuolas son vesículas constituidas por una membrana plasmática, y cuyo interior es predominantemente acuoso. Cuando en el contenido hay otro tipo de sustancias predominantes se habla de inclusiones.
Estructura de las vacuolas
Las vacuolas se forman a partir del retículo endoplasmático, del aparato de Golgi o de invaginaciones de la membrana citoplasmática. Las vacuolas de las células animales, que ya se han descrito al estudiar el RE y el AG, suelen ser pequeñas, y actualmente se denominan vesículas. Las vacuolas de las células vegetales suelen ser muy grandes, y sí reciben el nombre de vacuolas. Suele haber una o dos en cada célula. La membrana recibe el nombre de tonoplasto. Se forman mediante la unión de vesículas derivadas del RE y del AG, que suelen ocupar un 5% del volumen celular. A medida que la célula vegetal joven madura, las vacuolas crecen, llegando a ocupar un 50 %, y en ocasiones hasta un 90 %, de la célula vegetal madura. El conjunto de vacuolas de una célula vegetal recibe el nombre de vacuoma.
Funciones de las vacuolas
Las vacuolas poseen tres funciones principales:
- Acumular en su interior gran cantidad de agua. Con ello se consigue el aumento de volumen de la célula vegetal -turgencia celular- sin variar la cantidad de citosol o hialoplasma ni su salinidad. El agua, que realiza una función estructural, entra por ósmosis debido a la elevada concentración de sustancias que hay en las vesículas iniciales.
- Sirven de almacén de muchas sustancias. Unas son reservas energéticas elaboradas por la por la propia célula, como proteínas, otras son productos de desecho que resultarían perjudiciales si estuvieran en el citosol, otras son sustancias con funciones específicas, por ejemplo, los antocianósidos responsables de los colores de los pétalos, o los alcaloides venenosos para repeler los animales herbívoros, y otras son sustancias con función esquelética, como los cristales de carbonato cálcico y oxalato cálcico.
- Son medio de transporte entre orgánulos del sistema endomembranoso y entre éstos y el medio externo. Lo realizan las llamadas vesículas del RE y del AG.
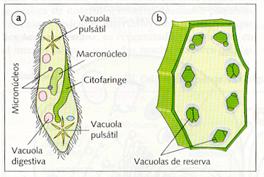
Por último, en las células animales se conocen dos tipos especiales de vacuolas: unas con función nutritiva, como las vacuolas fagocíticas y las pinocíticas, y otras con función reguladora de la presión osmótica; éstas son las vacuolas pulsátiles de los protozoos ciliados, que expulsan agua al exterior de una forma rápida, si la diferencia de presión es grande, o de una forma lenta, si los medios son isotónicos.
Entre las inclusiones, las más frecuentes son las inclusiones lipídicas, de aspecto muy refringente, que pueden contener lípidos de reserva o gotas de aceite, que por oxidación dan origen a las resinas y a los depósitos de látex, sustancia de la cual deriva el caucho natural.
a) Paramecio con vacuolas pulsátiles y digestivas.
b) Célula vegetal con vacuolas de reserva de sustancias oleosas.
|
|